三大营养物调节TAM功能
肿瘤相关巨噬细胞(TAM)占肿瘤质量的50%,是肿瘤微环境(TME)中最重要的免疫细胞,也是影响肿瘤进展、转移、复发的关键因素。其在TME中表现出强大可塑性,是具有肿瘤治疗的潜力靶点。
TAM的功能状态除受到各类生长因子信号调控外,还可通过代谢中的各类营养物质影响。
糖类
在糖代谢的整个过程中的多种产物参与调控TAM免疫抑制功能,与肿瘤发生发展关系密切。
糖酵解
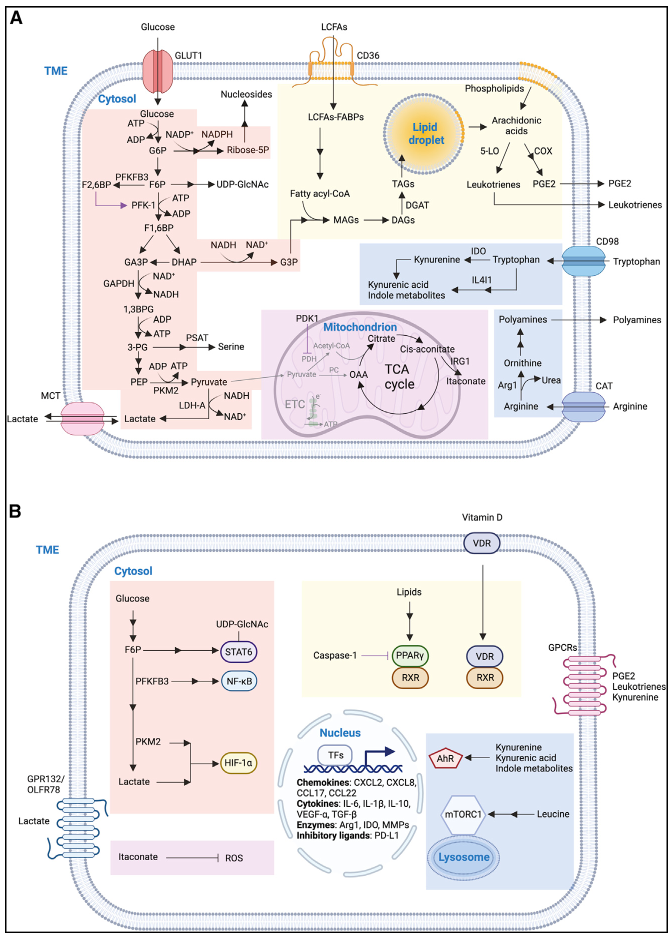
葡萄糖是机体主要的能量和碳来源,作为代谢原料通过葡萄糖转运蛋白(GLUT1)被TAM摄入,经过糖酵产生ATP和中间产物参与多种生物合成支持TAM代谢。在肿瘤发生时,TAM的免疫抑制功能依赖其高表达的GLUT1。
肿瘤发生时TAM糖酵解相关基因表达水平升高,参与调节TAM信号传导,并诱导趋化因子和PD-L1表达,促进TAM产生免疫抑制功能。(在人类HCC样本中就观察到了CD14+巨噬细胞在肿瘤相关细胞因子诱导下表现出高度的糖酵解活性。)
糖酵解产物为丙酮酸。
乳酸生成
由于肿瘤内部的缺氧环境,糖酵解生成的丙酮酸并不能正常进入TCA,而是被乳酸脱氢酶(LDH)还原为乳酸。人类和小鼠肿瘤模型中均观察到LDH-1表达增加。
同时,在缺氧环境中缺氧诱导因子HIF-1α通过诱导丙酮酸激酶(PDK1)磷酸化,使丙酮酸脱氢酶(PDH,催化丙酮酸生成乙酰辅酶A)失活,同样促进丙酮酸进入乳酸代谢途径。
在K-Ras介导的肺癌模型中,通过消耗LDH-A减少乳酸的产生,能够降低PD-L1和VEGF-α表达,抑制肿瘤进展。
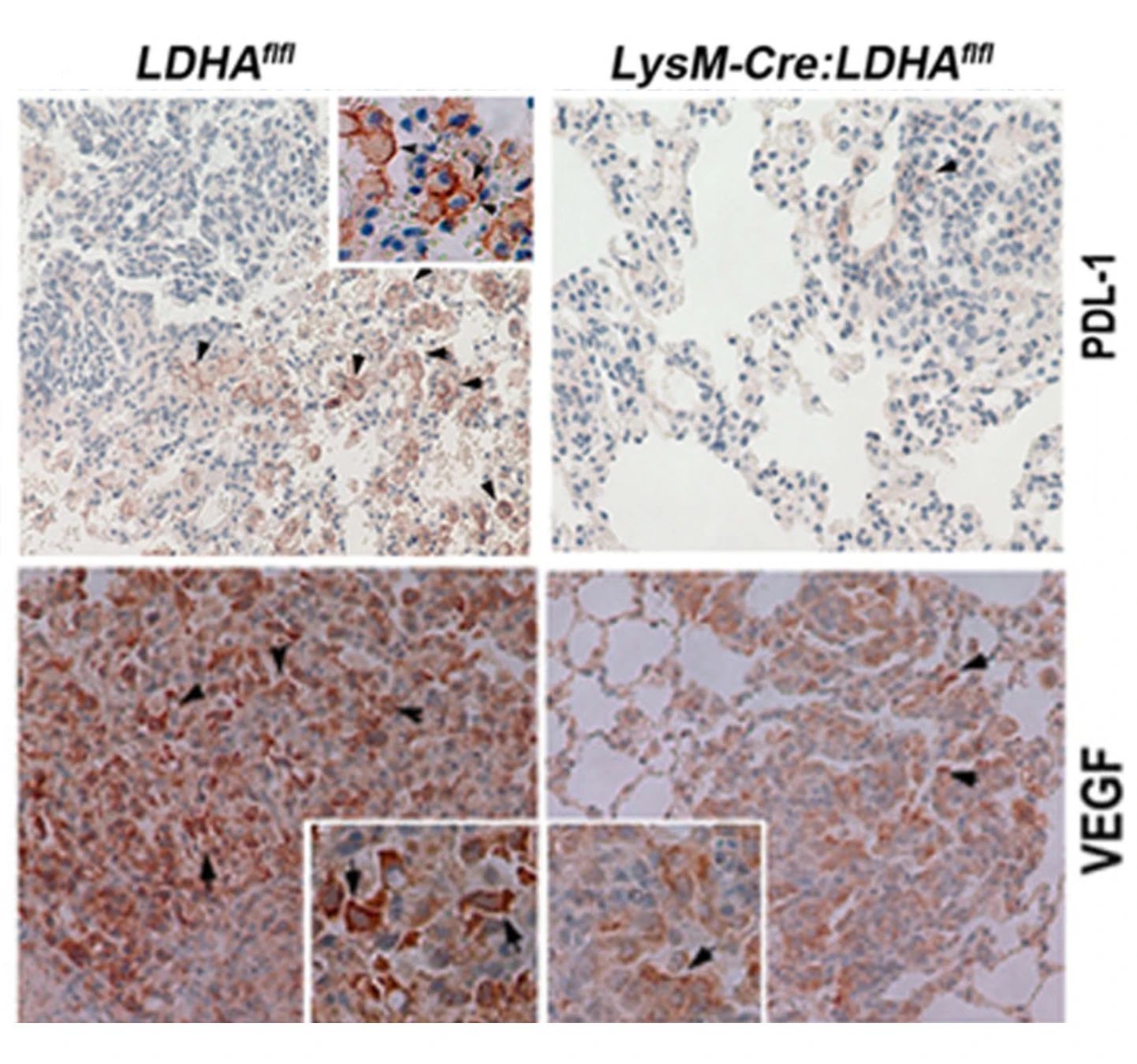
Cancer Res. 2017 Jul 1;77(13):3632-3643.
除作为代谢产物外,乳酸还是一类信号分子,在TAM膜上G蛋白偶联受体GPR132在感知乳酸信号后,能够促进多种趋化因子表达,并促进肿瘤转移、进展。肺部肿瘤上的气味受体Olfr78能够与GRP132协同感知乳酸信号,促进肿瘤生长、转移。
而在生成丙酮酸过程中,多种中间产物参与TAM代谢。
- 果糖6磷酸(F6P),通过己糖胺合成途径(HBP)的产物UDP-GlcNAc可作为糖基化反应的糖基供体,在乳腺肿瘤模型中与TAM免疫抑制功能相关,并且在高血糖条件下促进肿瘤进展。
- 磷酸二羟丙酮(DHAP),可被GPD1催化转化为甘油3磷酸(G3P),在GPAT进一步催化下生成磷脂酸,是三酰甘油(TAG)生物合成必需物。TAG代谢在TAM中具有重要功能(脂类)。
- 3磷酸甘油酸(3-PG),被PSAT催化下成丝氨酸,参与一碳代谢。在B16-F10肿瘤移植模型中,通过PSAT1耗竭能够减少抑制性TAM的数量,并同时抑制肿瘤生长。
三羧酸循环(TCA)
TCA是多种代谢的连接枢纽,通过产生NAD和FADH2参与电子传递链(ETC),进而由氧化磷酸化(OXPHOS)生成ATP。
在缺氧的TME中,TAM不能进行正常的ETC,通过TCA其他代谢中间物参与OXPHOS。
- 衣康酸,由IRG1催化顺乌头酸产生,可抑制琥珀酸脱氢酶(SDH)活性来防止线粒体产生ROS,从而发挥抗炎作用。在肿瘤接种后的模型中,IRG1及衣康酸表达升高,而敲除TAM中的IRG1能够抑制降低肿瘤负荷。
- 琥珀酸盐,由肿瘤细胞产生,通过激活TAM琥珀酸盐受体SUCNR1,由PI3K-HIF-1α通过触发促瘤信号。但同时琥珀酸盐还通过维持HIF-1α稳定促进TAM产生IL-1β,因此其在TAM中的功能还有待阐明。
脂类
TAM中脂滴形成见于多种人类和小鼠恶性肿瘤中,是由于在TEM缺氧条件下由线粒体驱动的脂肪酸氧化(FAO)水平降低,脂质降解减少造成。
脂肪酸摄取造成脂质积累
TAG是脂滴的主要成分,通过ATGL→HSL→MGLL酶系统被催化生成游离脂肪酸和甘油,TAM中MGLL表达降低,是脂质积累的一个原因。
相较于脂肪酸从头合成,通过摄取胞外脂肪酸才是维持TAM免疫抑制表型的重要因素。
CD36也称为脂肪酸转位酶,TAM可以通过CD36摄取长链脂肪酸(LCFA)在内的多种底物,是诱导其促瘤表型的关键。同时TAM可通过CD36摄取肿瘤微泡建立转移前生态位。
进入胞质的脂质可与过氧化物酶体增殖物激活受体(PPAR)结合,PPAR被激活后进入细胞核与RXR形成异二聚体,诱导脂质分解代谢基因表达。同时PPAR-γ与caspase-1结合后能够抑制FAO,同样造成脂质积累。
信号传导
脂质及其衍生物同样可发挥信号分子功能。
- PGE2,由花生四烯酸在环氧合酶(COX)和PGES催化下生成,与受体结合激活下游cAMP和Ca2+信号通路。花生四烯酸-PGE2通路基因表达与TREM2+TAM正相关。与T细胞共培养系统中,巨噬细胞产生的PGE2能够减弱CD4+T细胞增殖。
- 白三烯,同样由花生四烯酸在被5-LO催化下生成,发生肿瘤时TAM中5-LO表达降低,会抑制T细胞募集发挥免疫抑制功能。
- 维生素D,由胆固醇加工生成,与VDR、RXR相继结合形成异二聚体控制靶基因表达,抑制D-VDR结合过程能够抑制移植乳腺癌模型中的肿瘤进展,但目前具体机制不明。
- β-羟基丁酸(BHB),是酮体中的一类,能够抑制巨噬细胞中NLRP3炎性体引其的炎症反应。BHB也可作为信号分子,在与GPR109a结合后能够促进组织修复。
氨基酸
除作为蛋白质合成原料外,氨基酸同样可作为信号分子通过Rag-GTPase介导的营养感知通路参与mTORC1对TAM代谢的调节。
亮氨酸
亮氨酸可被TAM通过CD98摄取,CD98由SLC3A2和SLC7A5组成,通过抑制SLC7A5能够减少mTORC1介导的糖酵解和各类炎性细胞因子的产生。
色氨酸
色氨酸同样可被CD98摄入,巨噬细胞中的IDO1可以催化其生成犬尿氨酸,这个途径可以上调T细胞PD-1的表达水平,并且能够抑制体外T细胞增殖和活化。(而且巨噬细胞表达IDO还可促进其对凋亡细胞的”find me“信号的免疫耐受。推荐阅读:Efferocytosis:不同阶段分子差异与药物靶点)
有研究表明,IDO过表达的B16黑色素瘤具有免疫抑制表型,在一定程度上取决于犬尿氨酸-AhR介导的Treg-TAM相互作用。
此外,色氨酸被微生物群降解后产生含吲哚代谢物可以激活TAM中的AhR,通过抑制肿瘤内CD8+T细胞促进PDAC肿瘤进展。
IL4Ⅰ1是一种L-氨基酸氧化酶,是AhR通路的有效激活剂,能够促进色氨酸分解为吲哚代谢物和犬尿氨酸。富集IL4Ⅰ1+IDO1+PD-L1+TAM细胞群与多种人类肿瘤中T细胞障碍相关。【24】
除AhR外,犬尿氨酸还可与细胞表面受体GPR35结合,GPR35耗竭能够抑制遗传和致癌物诱导的CRC肿瘤进展。
精氨酸
精氨酸属于带正电荷氨基酸,可被阳离子氨基转运蛋白(CAT)SLC7A1-4吸收。进入TAM后被精氨酸酶1(Arg1)转化为尿素和鸟氨酸,参与鸟氨酸循环,最终产物腐胺可合成多胺参与细胞增殖。精氨酸-多胺途径在肿瘤浸润性骨髓细胞中被诱导并促进其存活,消耗多胺可以延长小鼠脑肿瘤抑制模型的存活率。
肿瘤发生时,在HIF-1α、乳酸、GM-CSF信号刺激下,TAM中高表达Arg1,并大量摄取精氨酸维持免疫抑制功能。
谷氨酸
谷氨酰胺可被谷氨酰胺酶(GLS)转化为谷氨酸,使用JHU083抑制GLS能够将来TAM重编程为促炎表型。谷氨酰胺-谷氨酸代谢可与葡萄糖代谢互作调节巨噬细胞反应。
参考资料
- Zhang X et al. Control of tumor-associated macrophage responses by nutrient acquisition and metabolism. Immunity. 2023 Jan 10;56(1):14-31.
- Vitale I et al. Macrophages and Metabolism in the Tumor Microenvironment. Cell Metab. 2019 Jul 2;30(1):36-50.
- Seth P et al. Deletion of Lactate Dehydrogenase-A in Myeloid Cells Triggers Antitumor Immunity. Cancer Res. 2017 Jul 1;77(13):3632-3643.